马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?立即注册

x
0 s- b; O( x* {/ o! j转载自:小药说药
' I; I' |; _1 p2 M) _; c) U0 i
+ k+ }* {! E# ^" {* W/ [: X/ m+ `' [前言 ! g! i* z8 T3 O+ V
自噬是一种调节机制,可去除不必要或功能失调的细胞成分,并回收代谢底物。针对肿瘤微环境中的应激信号,肿瘤细胞和免疫细胞中的自噬途径发生改变,从而对肿瘤进展、免疫和治疗产生不同影响。
1 G5 f7 G0 }7 j+ r* A! G% A8 e% D. P! J
最近的研究表明,自噬途径参与免疫细胞亚群的存活和凋亡、分化、激活、效应器功能以及向肿瘤的转运。同时,肿瘤自主自噬可以通过调节免疫反应改变肿瘤生长。此外,通过将免疫检查点疗法与自噬抑制剂相结合,可以消除自噬的促肿瘤作用。因此,自噬在癌症治疗中是一个复杂但很有前景的靶点。7 Q9 M; ]; m" x* M; ~
" {( p9 ?/ L# K8 O w
肿瘤细胞的自噬 , O: H" s: N$ q: x$ k& v" o+ _
肿瘤微环境(TME),在癌症进展、转移和治疗抵抗中起着关键作用。在TME中,肿瘤细胞中的自噬可由细胞内和细胞外的应激信号诱导,包括代谢应激、缺氧、氧化还原应激和免疫信号。* r# F* f' G; Z- O, ~2 O% h
* U% u n( E( S5 c9 l& I
代谢应激 & Z D* a8 D! I" `4 h2 ^
TME摄取的营养不足会影响代谢机制,导致细胞内代谢应激。为了应对代谢应激,肿瘤细胞通过上调营养转运体和激活自噬重新构建自己的代谢途径。
( Q1 z& ~9 \7 _
; Q! O$ u1 z4 @在机制上,5′-AMP活化蛋白激酶(AMPK)和mTOR复合物1(mTORC1)是两种相反的调节激酶,在营养缺乏的情况下改变自噬诱导。作为一种AMP传感器,AMPK通过AMP和ATP比率的增加而被激活,而mTORC1的活性由于氨基酸缺乏而降低。这导致自噬起始前复合物中靶点的磷酸化,从而启动自噬。
7 S& Q9 D2 d* w; w) U
+ B& p% l( c6 a) H2 p低氧应激
0 d% t, d& }1 h; ]1 ^& F缺氧是实体瘤的特征,缺氧导致线粒体氧化磷酸化受到抑制,AMP与ATP的比率增加,AMPK激活,缺氧同时也抑制mTOR信号。此外,缺氧导致激活转录因子4(ATF4)激活,从而使LC3B和自噬相关蛋白5(ATG5)表达上调,并维持高水平的自噬。
6 R/ u" ~* E/ e: R, v. D5 A- @0 j1 B) l* O' S
氧化应激
& y) u8 F2 J+ z8 a* N+ ]氧化应激反应了自由基和抗氧化剂之间的不平衡。活性氧(ROS)通过氧代谢和其他过程在细胞内产生,过量的活性氧可能增加DNA损伤的风险并促进肿瘤的发生。作为对ROS升高的反应,毛细血管扩张性共济失调突变(ATM)通过细胞质中的肝激酶B1(LKB1)和AMPK代谢途径激活TSC2肿瘤抑制因子,以抑制mTORC1并诱导自噬。氧化应激还可通过NF-κB介导的p62/SQSTM1上调促进自噬。
2 y. [4 n( P6 F+ {* z! w7 W5 e6 b' n H2 e
免疫信号 ; u' p9 D5 I7 h* _" ?
免疫信号可以调节TME中的自噬途径。损伤相关分子模式(DAMP)和细胞因子是调节自噬反应的主要介质。细胞外DAMP信号被细胞外或细胞内模式识别受体感知,如Toll样受体(TLR),当各种TLR识别DAMP并激活下游信号时,诱导自噬诱发生。
+ c0 @' J: W$ i. x$ e1 y: C1 l$ v0 M& F6 H: y! \, M9 W
此外,在果蝇中,细胞因子如TNF和IL-6样信号可以激活自噬,从而促进早期肿瘤生长和侵袭。TGF-β通过SMAD依赖和SMAD非依赖途径增加BECN1、ATG5和ATG7的转录水平,并激活自噬,在体外可以延缓人肝细胞癌和乳腺癌细胞的凋亡。
! u7 w8 W2 Y6 Y! Y% p) o4 V7 g: O* J! x& y
自噬介导的免疫逃避 ! E, j' D5 |" }1 A7 ~3 e& \0 X
逃避抗肿瘤免疫反应是各种肿瘤的重要生存策略。最近的证据表明,自噬在肿瘤免疫逃避中起着重要作用。有研究发现,胰腺导管腺癌(PDAC)中MHC I类分子的下调是通过选择性自噬降解介导的,抑制自噬释放出强烈的抗肿瘤免疫反应。+ ]3 U" o' Z! C" r N2 P
5 E; [6 w I, S p0 H
另一方面,MDSCs在TME中发挥免疫抑制作用,有研究证明MDSC中的自噬是抑制黑色素瘤抗肿瘤免疫活性的关键机制。MDSC免疫细胞中的自噬是降解MHC II类分子的中心,阻止抗肿瘤T细胞的启动和激活。& S" f8 p8 V* ], H' e
0 @ X; w8 i h- B. c* n
自噬与耐药性 - b# m9 a2 N$ u, ^; O
自噬作为(癌症)细胞应对威胁性应激源的机制,被认为是癌症治疗中治疗抵抗的重要机制。已有证据表明肿瘤细胞对顺铂的耐药性至少部分是由卵巢癌细胞系中自噬增加介导的。类似的证据表明顺铂、阿霉素和甲氨蝶呤通过抑制骨肉瘤中的自噬来克服化疗耐药性。0 f _' i* c: y9 ?: m- [% Q
0 V- G6 P Z; ~+ K& _2 ]! c$ u% e
有趣的是,例如顺铂和自噬之间的相互作用是一个连续统一体,甚至在体外和体内对某些化疗药物本身不具有耐药性的细胞中,添加自噬抑制剂,如氯喹(CQ)也可以提高治疗效果。这已在肾上腺皮质癌的小鼠模型、结肠癌细胞系和5-氟尿嘧啶以及替莫唑胺诱导的胶质瘤细胞的细胞毒性中得到证实。8 b. n. b b# W$ [% \$ U- Z
2 F' }# k' m3 [; `5 X
除此之外,对基于抗体的治疗也进行了类似的结果,例如在曲妥珠单抗耐药的乳腺癌细胞中,使用CQ的自噬抑制导致肿瘤的几乎完全消除。同样,使用CQ抑制自噬也能够有效对抗贝伐单抗诱导的结直肠癌细胞自噬,减少体内小鼠肿瘤模型中的肿瘤生长。7 y; J6 I D( J
/ r! z7 X7 I6 T$ b+ y1 Q1 j( W- s
自噬在肿瘤治疗中的研究现状 ' A3 J$ ]7 R* }
自噬抑制剂分为针对ULK1/ULK2或VPS34的早期抑制剂,如SBI-0206965、3MA和wortmannin,以及针对溶酶体的晚期抑制剂,如CQ、羟基氯喹(HCQ)、巴非霉素A1和莫能菌素,CQ和HCQ通过干扰溶酶体酸化抑制自噬体降解。然而,在临床试验中,HCQ单一疗法未能控制晚期胰腺癌患者的肿瘤生长,目前多将自噬抑制与其他癌症治疗相结合,以提高治疗效果。
4 D' d/ y1 S# F8 z7 U8 _2 x0 S! ?3 D* D6 n6 _ P5 M' O
化疗 9 {7 R/ o. H! _- z
癌症中的高自噬通量与化疗反应降低相关,并与癌症患者的低生存率相关。临床前研究表明,抑制自噬可以克服NSCLC、膀胱癌、甲状腺癌和胰腺癌的化疗耐药性。此外,一些研究的结果表明,自噬抑制可能与MEK–ERK信号的抑制产生协同作用。% D; _, R3 g/ \3 c& B9 M |
' B. R; |; g7 q# {2 r+ Y: d- ]" n
2014年的一项早期II期研究使用HCQ单一疗法治疗转移性胰腺癌患者,这些患者以前曾通过其他方法治疗,主要终点为两个月的无进展生存期。结果,不同患者的自噬水平不同程度地降低,但主要终点没有显著改善。另一项联合HCQ、吉西他滨和nab紫杉醇治疗晚期或转移性胰腺癌患者的研究也未能证明延长12个月的总生存期。然而,重要的是,采用HCQ的患者显示出了显著且更好的应答率(38.2%对21.1%)。
& U+ N+ M7 v/ h8 K
4 h. n" v. X4 Q' N; y放疗 8 ~* b `# M X, _+ L0 j
自噬在保护肿瘤细胞免受放射治疗引起的细胞死亡方面起着关键作用。在乳腺癌细胞中,辐射诱导自噬相关基因的表达,伴随自噬体的积累。在放疗的同时短期抑制自噬可增强放疗对耐药癌细胞的细胞毒性。同样,低氧通过诱导自噬增强A549肺癌细胞的放射抗性。
; n6 h9 R- D4 E% I- s. Z( i8 O) T" x# @3 N( W
在胶质母细胞瘤中,放射治疗通过增加哺乳动物STE20样蛋白激酶4(MST4)的表达诱导自噬,该蛋白激酶通过ATG4B磷酸化来刺激自噬。小分子抑制剂NSC185058(靶向ATG4B)与放疗联合使用,可损害胶质母细胞瘤的颅内异种移植生长,并延长治疗小鼠的生存期。因此,靶向肿瘤自噬可能增强放射治疗的疗效。事实上,在癌症患者的临床试验中,已经利用了自噬抑制剂与放射治疗相结合。
3 S( i: b9 U* Z2 U, s4 ~* m$ k, \) I6 v) J
免疫疗法 ; ]9 j, b* a Q$ D2 i+ q
利用免疫系统是对抗癌症的重要途径。抑制自噬可能会损害系统免疫,因为自噬涉及免疫系统发育和效应T细胞的存活和功能。然而,在黑色素瘤和乳腺癌的临床前模型中,CQ在短时间内对自噬的系统性抑制并没有损害T细胞功能。数据表明,免疫系统可能对某种程度的自噬抑制具有耐受性。然而,鉴于自噬可以调节肿瘤免疫反应,靶向自噬可以提高免疫治疗的疗效,克服免疫治疗的耐药性。/ r, W. g5 a: Z) d0 {* y0 U
# ]6 _# Y3 v. V+ O2 C例如,使用抑制剂SB02024或SAR405抑制VPS34激酶活性导致TME中CCL5、CXCL10和IFN-γ水平升高,从而使黑色素瘤和结直肠癌模型中NK细胞和T细胞肿瘤浸润水平升高。在这些模型中,VPS34抑制也逆转了抗PD1或抗PD-L1治疗的耐药性。此外,CQ治疗可阻断自噬介导的MHC I类降解,与双重ICB治疗(抗PD1和抗CTLA4抗体)协同作用,在胰腺癌小鼠模型中产生增强的抗肿瘤免疫反应。4 B( e X$ V8 F2 r" C4 G; P
2 a# A4 `: a, @6 \6 ]
因此,靶向自噬可能增强免疫治疗。目前正在进行HCQ联合免疫治疗治疗不同类型癌症患者的临床试验。
5 h2 o' z; v! R4 y1 d" a! z0 ^$ I3 ]# j) U3 y
此外, CAR-T细胞疗法在治疗血液肿瘤方面取得了临床成功,但在治疗实体癌方面依然疗效有限。自噬调节可能为使用CAR-T细胞治疗的癌症患者提供一些益处。众所周知,TME是实体瘤中CAR-T细胞浸润和功能的屏障,鉴于自噬抑制可重塑TME并促进TH1型趋化因子的产生,自噬抑制可促进CAR-T细胞向肿瘤的转运,CAR-T细胞的自噬增强可能支持T细胞在TME中的适应性和存活。此外,肿瘤自噬抑制可能导致抗原表达增加,从而增强CAR-T细胞介导的肿瘤杀伤作用。最后,自噬抑制可能改善细胞因子释放综合征并为患者带来临床益处。总的来说,抑制自噬以提高免疫治疗效果的潜力是一个不断探索的有希望的领域。
& f8 j+ ?" ?- L
8 K7 E6 z# G$ c$ G9 ]# x% k! z/ H7 |小结 1 b. a. c- B4 D- t
噬是肿瘤生物学、免疫学等多个领域研究的重要机制。鉴于TME中不同细胞中的不同因子可诱导自噬,其诱导和激活可促进或抑制肿瘤进展。T细胞亚群中的自噬可能在抗肿瘤免疫反应中发挥积极作用,而肿瘤细胞中的功能性自噬可能支持肿瘤抗原呈递和识别,在这种情况下,抑制自噬可能有害于抗肿瘤免疫。另一方面,自噬途径可能与肿瘤细胞存活、肿瘤抗原降解、TH1型趋化因子减少、Treg细胞和MDSC增强有关。因此,以自噬为系统靶向的癌症治疗方法具有挑战性。
" E8 N: E+ |1 A4 P) q+ u) u
4 P) F! M% C! N/ i8 p4 ^, {# m% V因此,为了在肿瘤免疫治疗中靶向自噬途径,探索主要免疫细胞亚群中自噬的生物学活性至关重要。最近的研究已经开始评估T细胞、巨噬细胞和树突状细胞的自噬途径。未来的工作可能会扩展到B细胞、NK细胞和NKT细胞。此外,随着癌症的进展,自噬途径会随着TME中不同刺激的反应而动态改变,必须了解自噬如何同时参与TME中免疫细胞、肿瘤细胞和基质细胞的功能和存活的机制,这样才能够最终使癌症患者获得临床益处。1 |1 ~" \2 _7 \3 |% H
; G) z# {0 p* O s) X参考文献
$ l7 G, ^# [3 h, p% m ^ 1.Autophagy in tumour immunity and therapy. NatRev Cancer. 2021 May; 21(5): 281–297.
5 g9 L, S+ }& j2 n8 i- d; ]0 N5 g2 C) {& v: Q6 X
2. Autophagy in Cancer Therapy-Molecular Mechanisms and Current Clinical Advances. Cancers (Basel). 2021 Nov8;13(21):5575.
7 i0 v N: G( H" D8 R5 x; F& }9 ?) K+ B1 w* }, l `: k, C
|
 完成手术,病理也出来了,烦请各位老
之前因为心脏问题,去心内造影,前降支75%狭窄,回到心胸外科手术,目前主治医生说是
完成手术,病理也出来了,烦请各位老
之前因为心脏问题,去心内造影,前降支75%狭窄,回到心胸外科手术,目前主治医生说是
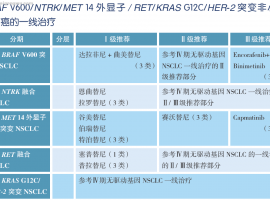 CSCO/NCCN双指南推荐!MET突变肺癌患
你知道吗?人体内存在一种掌控着生命密码的特殊蛋白——MET蛋白。正常的MET蛋白如同精
CSCO/NCCN双指南推荐!MET突变肺癌患
你知道吗?人体内存在一种掌控着生命密码的特殊蛋白——MET蛋白。正常的MET蛋白如同精
 求助: 阿美耐药, 化疗效果不好,身体
2022年7月确诊 右肺上叶近肺门肿块 纵隔淋巴结肿大 左侧锁骨淋巴结转移
求助: 阿美耐药, 化疗效果不好,身体
2022年7月确诊 右肺上叶近肺门肿块 纵隔淋巴结肿大 左侧锁骨淋巴结转移
 求助贴,奥西替尼一年多后CT有进展,
我父亲今年79,23年12月穿刺病理诊断为肺腺癌晚期。
基因检测EGRF19外显因子突变,丰
求助贴,奥西替尼一年多后CT有进展,
我父亲今年79,23年12月穿刺病理诊断为肺腺癌晚期。
基因检测EGRF19外显因子突变,丰
 EGFR靶点病人,到底适不适合免疫治疗
一直一直在听说,肺癌EGFR突变的病人,免疫治疗很难获益,甚至有超进展可能,具体哪里
EGFR靶点病人,到底适不适合免疫治疗
一直一直在听说,肺癌EGFR突变的病人,免疫治疗很难获益,甚至有超进展可能,具体哪里






 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡